杂种优势(Heterosis)是指F₁杂交后代在形态、生理或化学性状上优于其亲本的生物学现象,在自然界中广泛存在。查尔斯·达尔文在约150年前,首次对植物的杂种优势进行了科学而系统的描述[1]。在作物育种中,美国育种家约于上世纪30年代开始利用玉米的杂种优势[2],所选育的杂交品种产量可达自交系的近六倍。我国杂交水稻育种始于1964年,其杂交种相较于常规品种可实现20%–30%的增产[3]。
为解释杂种优势的机理,遗传学家先后提出了多种模型,包括显性(及假超显性)[4,5]、超显性[6-8]和上位性模型[9,10],分别强调有害等位基因的互补、位点杂合性以及位点间的互作。尽管分子生物学技术的发展推动了杂种优势遗传基础的研究,其分子机制至今尚未完全阐明。
近日,黄文超课题组在《Advanced Genetics》上发表了题为“Quantitative Insights into the Genetic Mechanisms of Crop Heterosis”的综述文章。该文从数量遗传学角度,系统总结了表型、转录、代谢和蛋白水平上不同遗传模式对作物杂种优势的贡献。文章指出,加性效应作为一种主效遗传模式,可用于预测F₁的杂种优势,从而加速杂交作物育种进程。

早在20世纪初,East便指出“杂种优势问题本质上是数量性状的遗传问题”(“the problem of heterosis is the problem of the inheritance of quantitative characters”)[11]。这里所说的“数量性状”,包括可定量测量的表型或分子表达水平。根据F₁与亲本之间的数量关系,遗传模式大致可分为四类(图1)[12]:加性(F₁等于双亲均值)、部分显性(F₁偏向某一亲本但未达到其值,且显著偏离双亲均值)、显性(F₁与某一亲本无显著差异)以及超显性(F₁优于高值亲本或劣于低值亲本)。
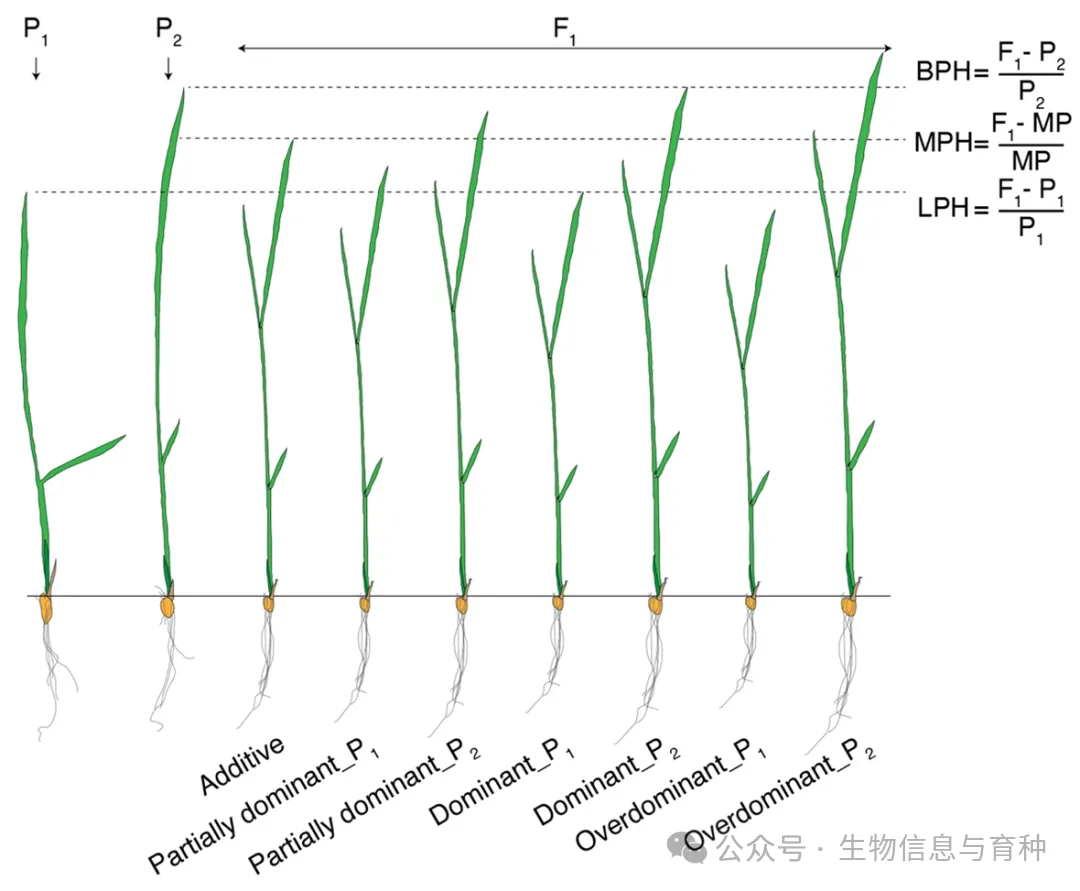
图1. 四种不同的遗传模式。
杂种优势源于杂交,而恰当的杂交设计对于准确评估遗传效应至关重要。目前,多数杂种优势研究仍基于较小规模的杂交群体(通常不超过10个F₁杂交后代)。针对大规模杂交群体的设计主要包括完全/不完全双列杂交(图2)、回交、测交、随机杂交、组间杂交、阶乘杂交和三向/四向杂交等。其中,完全双列杂交要求较高,需所有亲本可育且彼此之间均进行正反杂交。然而,该设计在全面解析杂种优势方面具有明显优势:它可以保证每个亲本在杂交群体中具有同等代表性,避免因后代数量不均而导致的评估偏差,且其均衡的群体结构也有助于准确评估父本和母本对F₁的贡献。
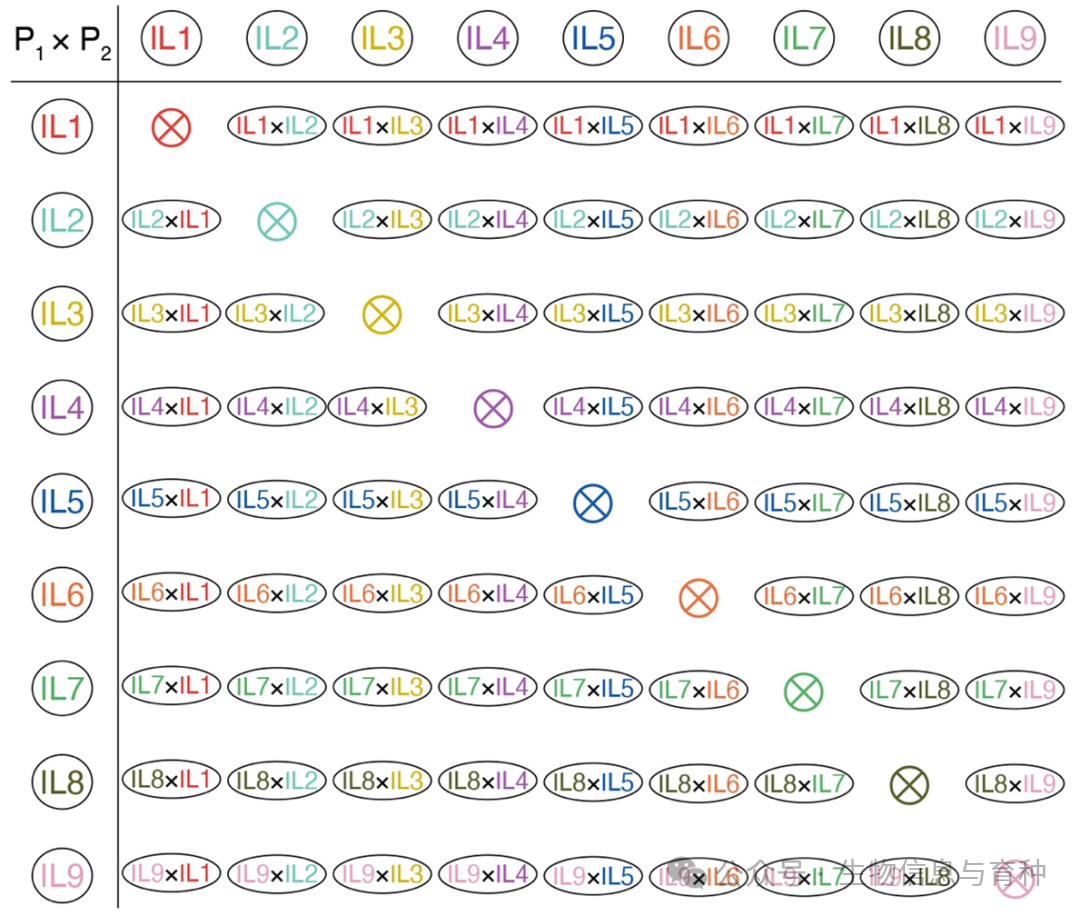
图2. 完全双列杂交设计。
在作物农艺性状分析中,可针对成分性状或复合性状分别计算杂种优势。复合性状(如产量、株高)由两个及以上成分性状(如粒重、粒数、节间数、节间长度等)构成。即使亲本的复合性状表现相似,只要其成分性状存在互补性差异,F₁仍可能表现出显著杂种优势,此现象称为“模拟显性”[9]。成分性状间的加性效应(包括加性×加性互作)是复合性状产生正向杂种优势的稳定来源,而维持成分性状间的适度平衡对实现杂种优势至关重要。
不同遗传模式对农艺性状杂种优势的贡献已在多种作物中广泛报道,但其贡献比例而异。例如,在玉米和水稻中,表型水平的加性、部分显性、显性、超显性效应以及多种上位性互作均被证实与杂种优势相关。转录组分析表明,加性表达模式在玉米和水稻中普遍存在,且常为基因表达的主要模式,这为理解杂种优势提供了重要线索。蛋白质组与代谢组研究,同样揭示了加性效应在杂种优势中的重要作用。
该综述提出了一个定量模型(图3),将不同遗传模式与复合性状(以水稻产量为例,简化为粒重与粒数两个成分性状)的杂种优势相联系。模型表明:当双亲产量相同时,成分性状间的加性×加性互作总是使F₁产生正向杂种优势;当涉及部分显性、显性或超显性效应时,F₁可能表现出不同方向与比例的杂种优势,其中极高正向优势的出现概率较低,而在超显性效应下出现严重负向优势的可能性更高。结合表型、转录、蛋白和代谢水平的结论,该模型同样适用于解释分子水平不同遗传模式对复合性状杂种优势的贡献。
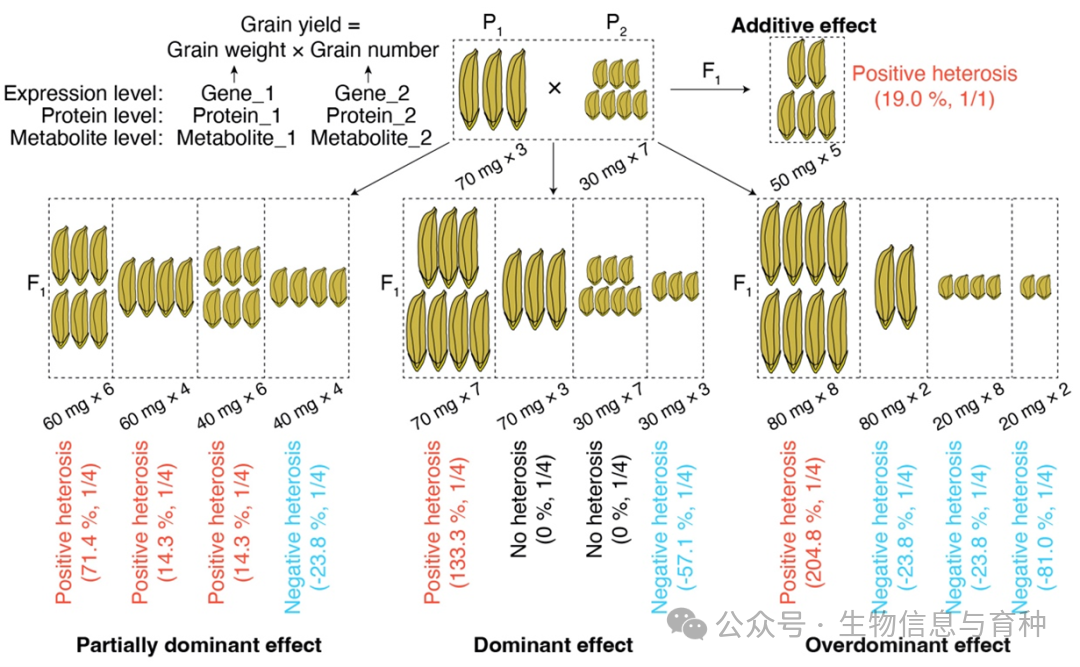
图3. 复合性状杂种优势的数量模型。
基因组下游分子(如转录本、蛋白质和代谢分子)是连接基因组信息与农艺性状的桥梁,其与表型的关联往往比基因组序列变异更为直接。在玉米和水稻中,代谢分子已成功被用于预测F₁表型或杂种优势,在某些情况下其预测准确度甚至高于基因组数据。在玉米中,转录组数据对部分性状杂种优势的预测能力优于基因组和代谢组。值得注意的是,转录或代谢水平的加性效应可准确预测水稻和玉米F₁的杂种优势。
总之,阐明作物杂种优势的分子机制仍是植物遗传学的重要挑战。尽管已提出多种遗传模型,但由于基因型、性状、组织、环境、发育时期、组学数据类型、物种及分析策略等方面的差异,研究结论常不一致。基于完全双列杂交设计,系统研究基因组下游分子的遗传模式,有助于理解杂种优势机理。虽然加性及其他遗传效应可解释相当比例的杂种优势变异,未来仍需开发更稳健的预测模型。这类模型的建立有赖于代表性亲本的选择、合适的杂交设计、复合性状的精细解析、多组织多组学数据的获取、时间序列分析、局部环境因素的考量以及恰当统计方法的综合运用。展望未来,结合环境与表观遗传学发现,对基因组下游分子进行定量分析,将深化对作物杂种优势的理解,加速优良杂交品种选育,从而为全球粮食安全作出贡献。
参考文献
[1] C. R. Darwin, The effects of cross and self fertilization in the vegetable kingdom, (1876).
[2] D. N. Duvick, “Biotechnology in the 1930s: the development of hybrid maize,” Nature Reviews Genetics 2 (2001): 69-74, https://doi.org/10.1038/35047587
[3] S. Lin, L. Yuan, “Hybrid rice breeding in China,” Innovative approaches to rice breeding International Rice Research Institute, Los Banos, Philippines (1980): 35-51,
[4] C. B. Davenport, “Degeneration, albinism and inbreeding,” Science 28 (1908): 454-455, https://doi.org/10.1126/science.28.718.454-b
[5] D. F. Jones, “Dominance of linked factors as a means of accounting for heterosis,” Proceedings of the National Academy of Sciences of the United States of America 3 (1917): 310-312, https://doi.org/https://doi.org/10.1073/pnas.3.4.310
[6] G. H. Shull, “The composition of a field of maize,” Journal of Heredity 4 (1908): 296-301, https://doi.org/10.1093/jhered/os-4.1.296
[7] G. H. Shull, “The genotypes of maize,” American Naturalist 45 (1911): 234-252, https://doi.org/10.1086/279207
[8] E. M. East, “The distinction between development and heredity in inbreeding,” American Naturalist 43 (1909): 173-181, https://doi.org/https://doi.org/10.1086/279042
[9] F. D. Richey, “Mock-dominance and hybrid vigor,” Science 96 (1942): 280-281, https://doi.org/10.1126/science.96.2490.280
[10] L. Powers, “An expansion of Jones's theory for the explanation of heterosis,” American Naturalist 78 (1944): 275-280, https://doi.org/10.1086/281199
[11] E. M. East, “Heterosis,” Genetics 21 (1936): 375-397, https://doi.org/10.1093/genetics/21.4.375
[12] Z. J. Chen, “Molecular mechanisms of polyploidy and hybrid vigor,” Trends in Plant Science 15 (2010): 57-71, https://doi.org/10.1016/j.tplants.2009.12.003
论文信息
Quantitative Insights into the Genetic Mechanisms of Crop Heterosis
Zhiwu Dan, Yunping Chen, Wenchao Huang
Advanced Genetics
DOI:10.1002/ggn2.202500030
原文链接
https://advanced.onlinelibrary.wiley.com/doi/10.1002/ggn2.202500030
欢迎加入生信AI育种交流群(一群已满,请加二群,入群请备注“姓名-单位”,方便交流)~~~